
Imageomics
Imageomics Future and Emerging Technologies (FET-OPEN) 2020: High-throughput imaging of proteins using nanobodies Horizon 2020 01.07.2021-31.12.2024 The conventional detection of proteins by imaging employs fluorophores


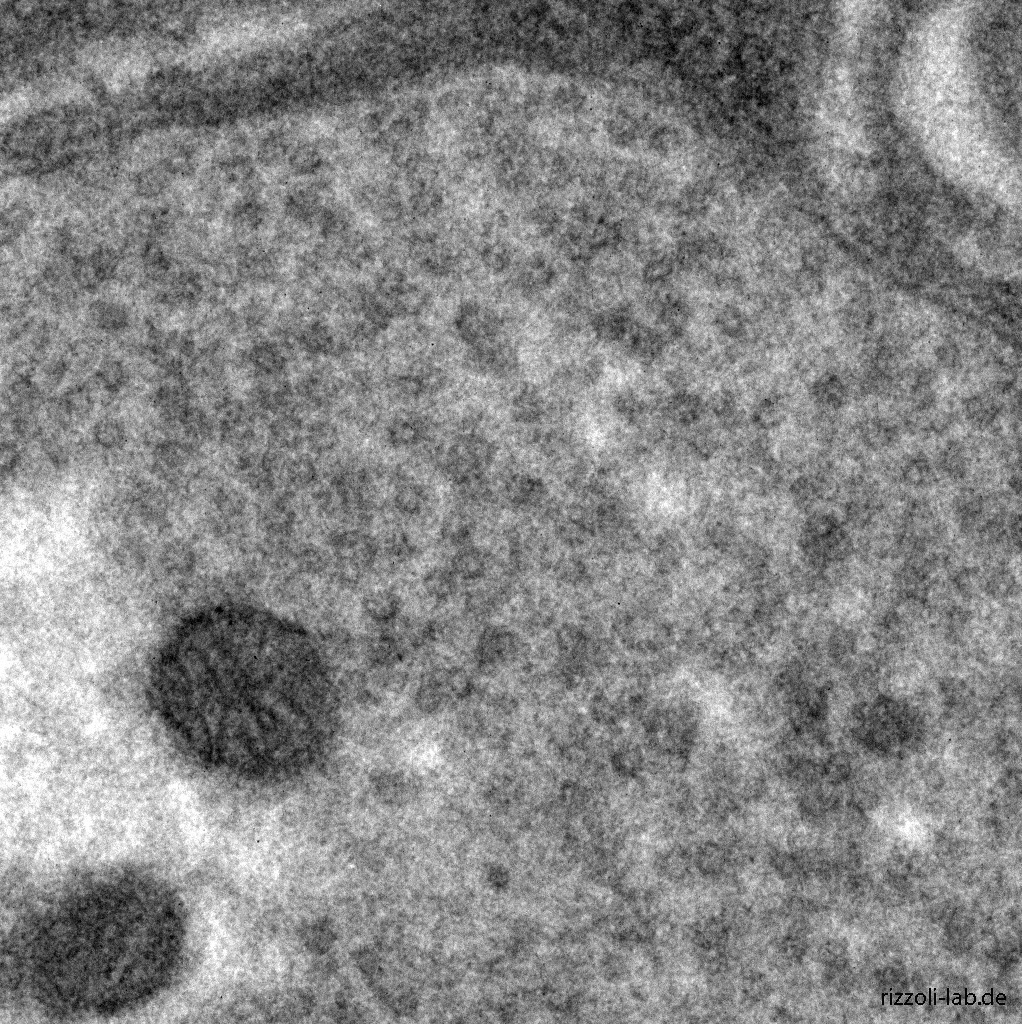
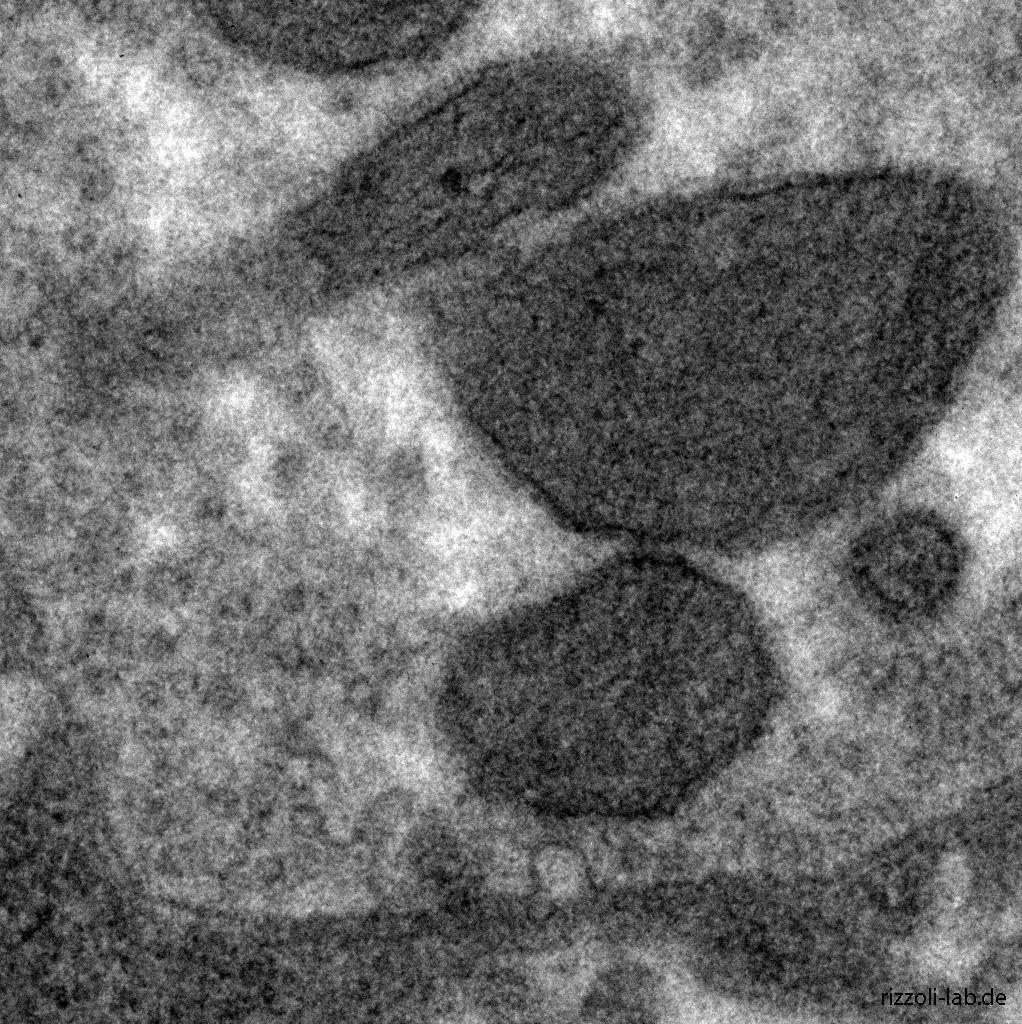
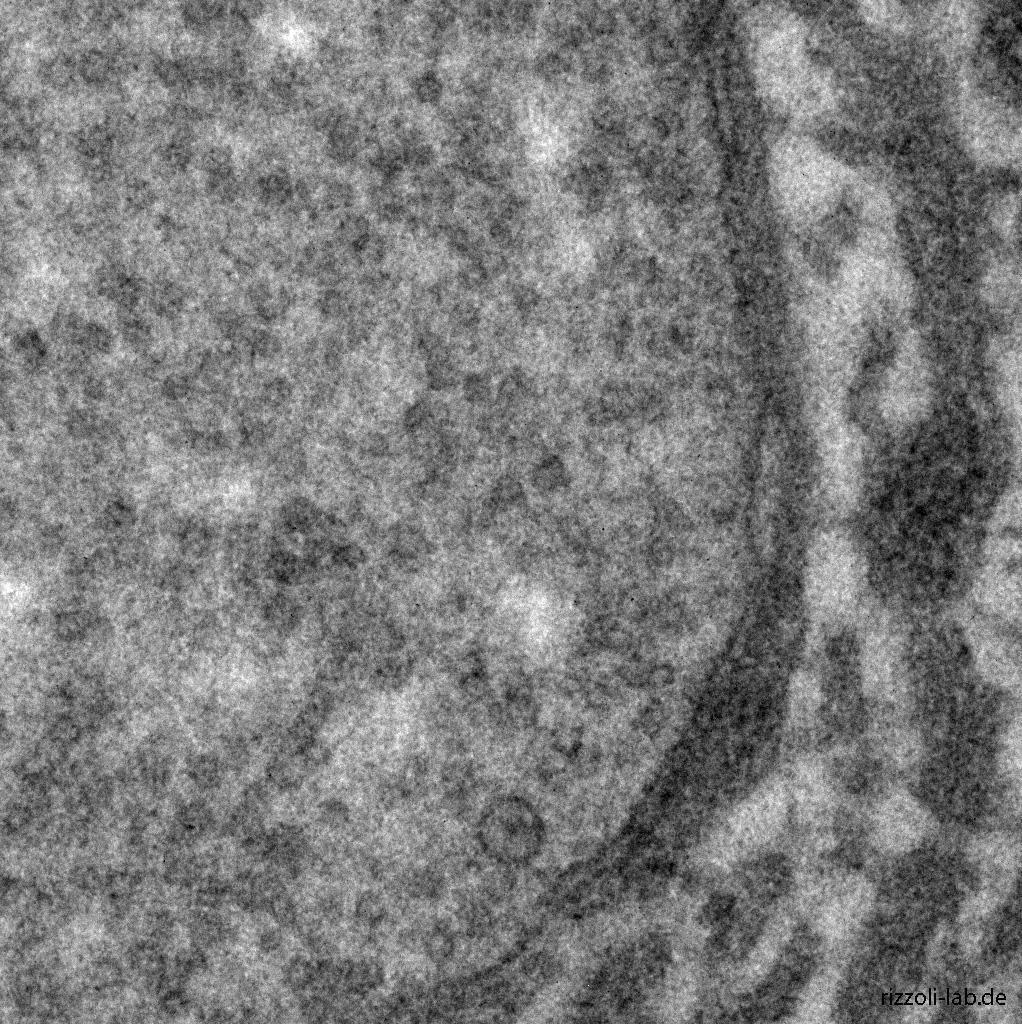
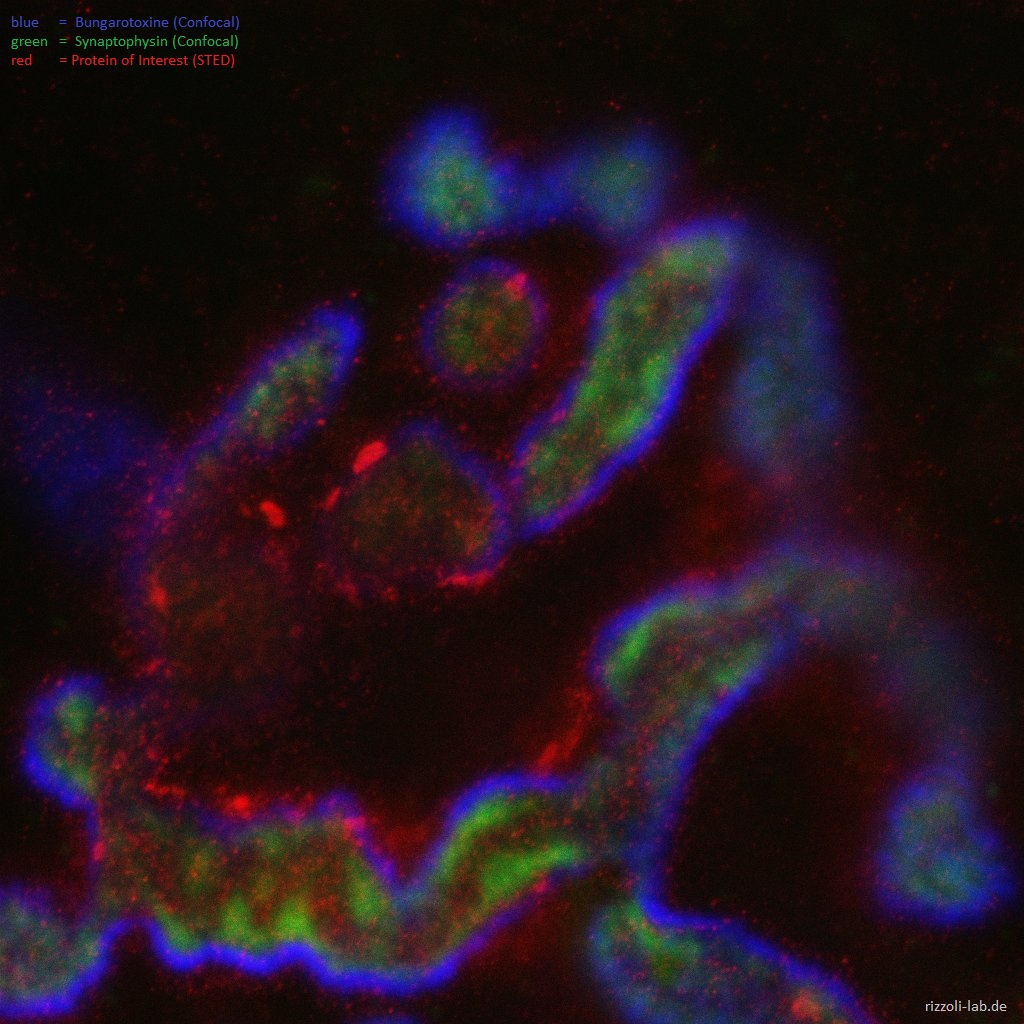
Imageomics Future and Emerging Technologies (FET-OPEN) 2020: High-throughput imaging of proteins using nanobodies Horizon 2020 01.07.2021-31.12.2024 The conventional detection of proteins by imaging employs fluorophores
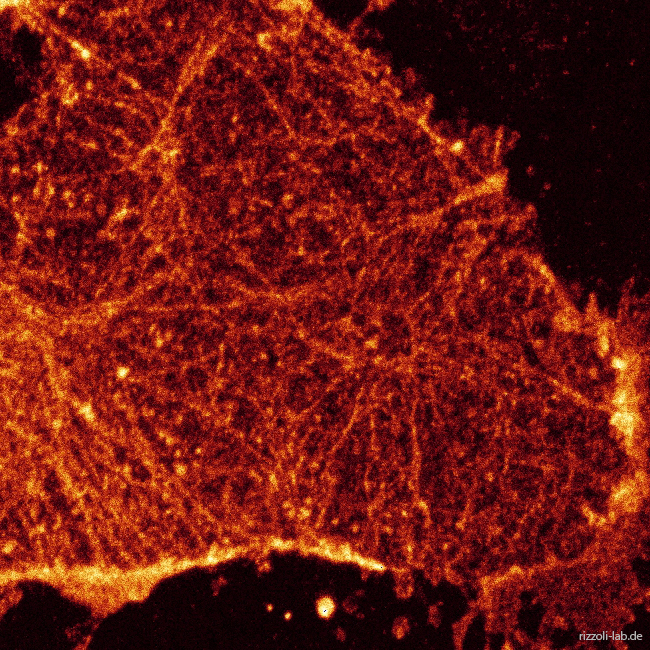
ONE Microscopy Optimized Nanoscale Expansion microscopy Super-resolution imaging has been applied to biological specimens for more than two decades. Nevertheless, it still falls short of

NanoSIMS 50L Starting at the end of 2017, the Center for Biostructural Imaging of Neurodegeneration (BIN) can offer the services of a new high performance

ULTRARESOLUTION Beyond super-resolution: ultra-resolution imaging provides solutions for synapse physiology and brain pathology ERC Synergy Grant 2020 01.06.2021-31.05.2027 Neurons contain hundreds of specialized proteins, whose
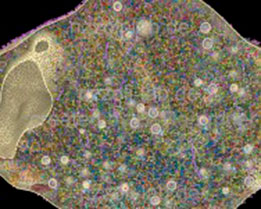
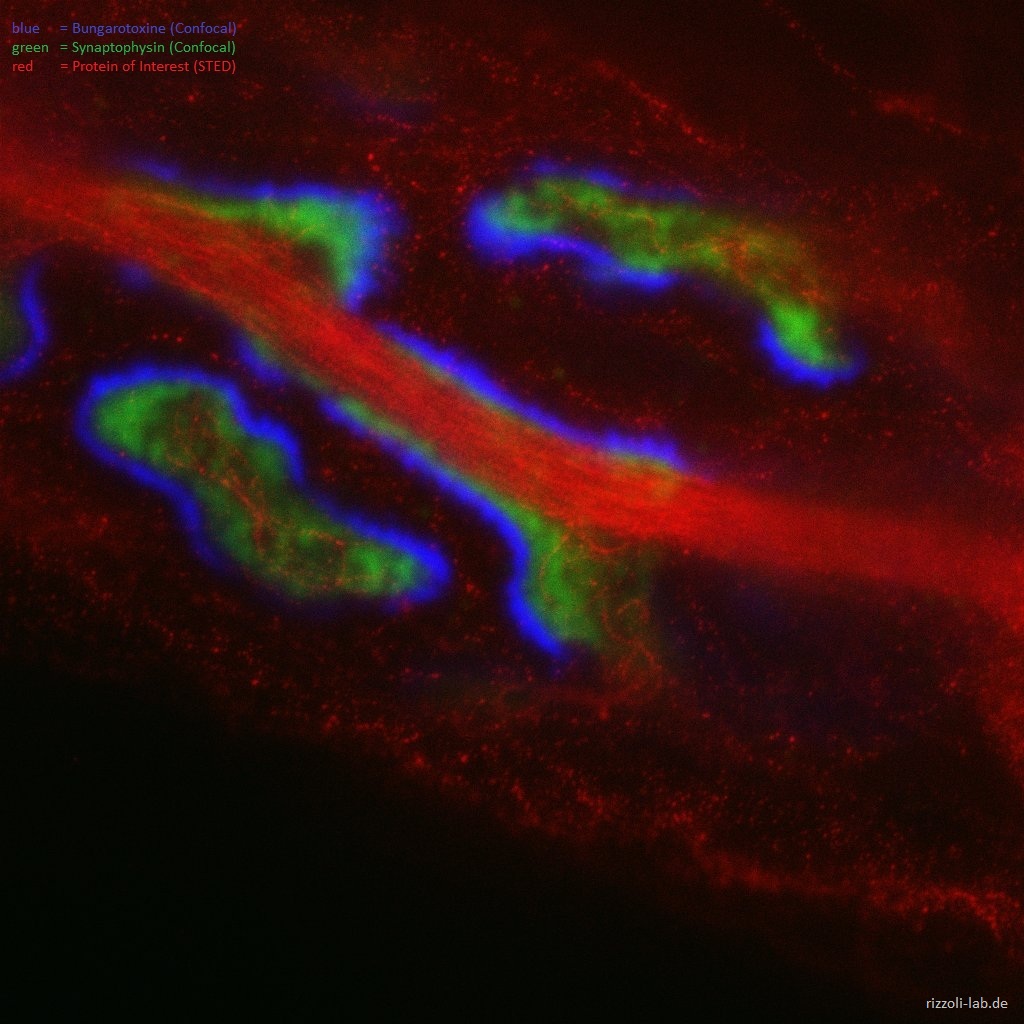
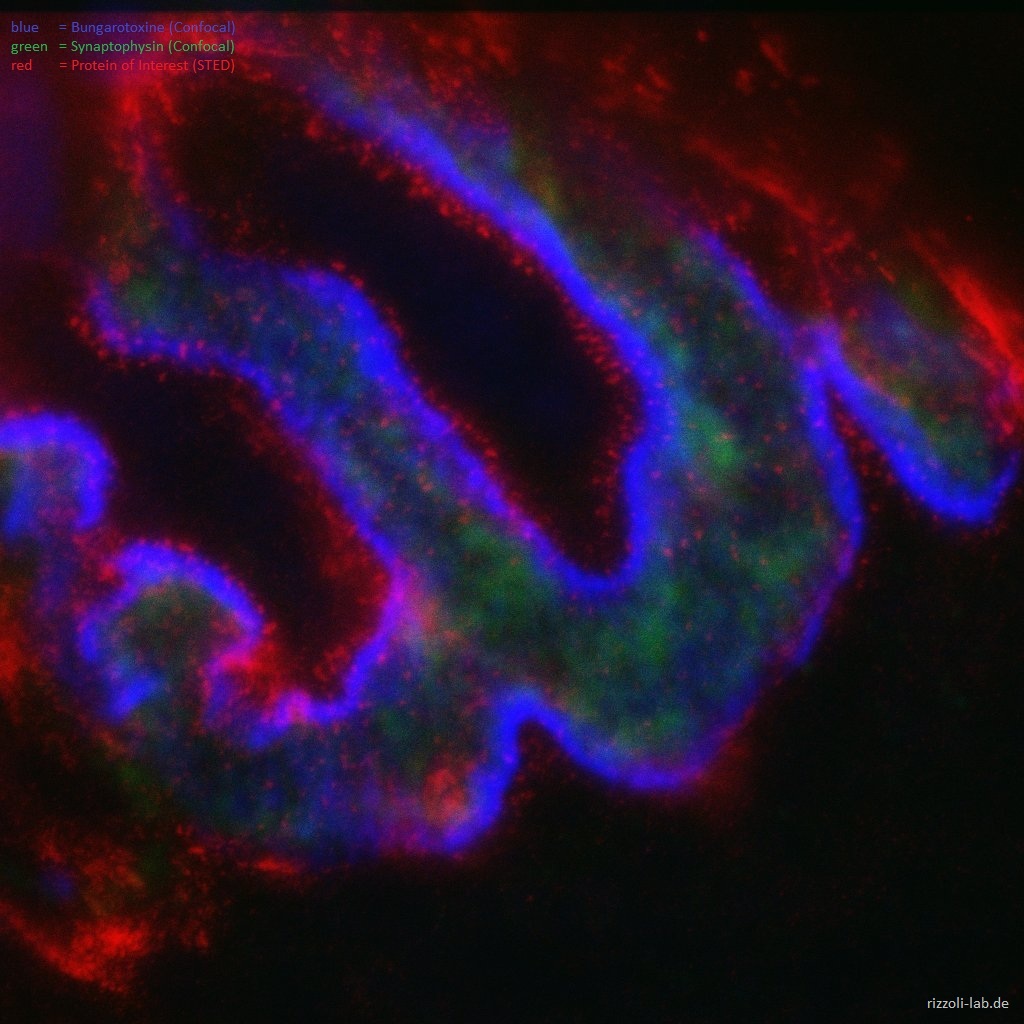
The Neuron Nanomap In this project, which has started in April 2014, we generate a super-resolution image – a nanomap – of presynaptic nerve terminals,